
|

"Portanto, o consórcio tomate+coentro e a irrigação por aspersão afetaram a distribuição espacial de adultos de mosca-branca e a incidência de viroses associadas. Esses resultados reforçam a importância do plantio do coentro antes do transplante do tomateiro para o campo" https://www.alice.cnptia.embrapa.br/handle/doc/657043
Finalmente, a análise de sequenciamento de RNA indicou que o CaLecRK-S.5 regula positivamente a imunidade da planta no nível transcricional. Em conjunto, estes resultados sugerem que a resistência de largo espectro mediada por CaLecRK-S.5 está associada à regulação do priming. https://academic.oup.com/jxb/article-abstract/67/19/5725/2236380 e
L-type lectin receptor kinases: New forces in plant immunity
divided over 4 clades (Fig 1) [6,7]. These 4 are also found in Nicotiana benthamiana and are
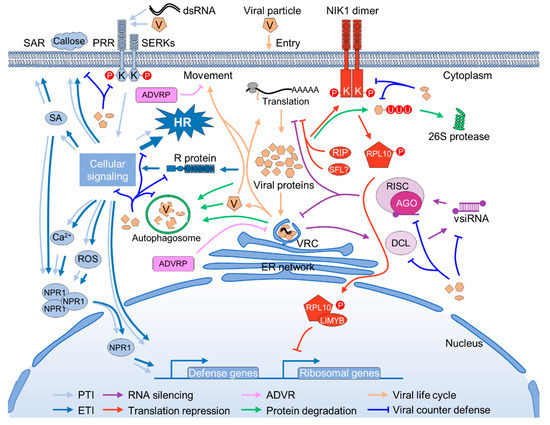
likely Solanaceae-specific [6]. In the more distant monocots, the clade composition is …Para deter a invasão de patógenos, as plantas - diferentemente dos animais - dependem exclusivamente de um sistema imunológico inato. Eles possuem um arsenal expandido de receptores de reconhecimento de padrões localizados na superfície celular (PRRs) para perceber padrões moleculares associados a danos e micróbios (M / DAMPs), aqui chamados coletivamente de padrões de invasão [1]. Exemplos proeminentes são a detecção de flagelina 2 (FLS2) e o receptor do fator de alongamento Tu (EF-Tu) (EFR), 2 quinases receptoras (RLKs) com ectodomínios de repetição ricos em leucina (LRR) que iniciam a defesa após o reconhecimento de flagelina bacteriana e EF -Tu, respectivamente [2]. Os principais atores emergentes na imunidade de plantas são as quinases receptoras de lectina, RLKs que são subdivididas em 3 classes distintas baseadas em seus domínios extracelulares de lectina, isto é, G- (relacionado a GNA ou S-locus), C- (dependente de cálcio) e L- (leguminosa) tipo [3]. Todos os 3 são onipresentes em plantas, mas ausentes em animais, que implantam receptores distintos de lectina do tipo C (CLRs) para iniciar a imunidade inata [4]. Os últimos anos testemunharam um interesse acelerado em quinases de receptores de lectina de plantas. Nesta pérola, resumimos nosso conhecimento atual sobre as quinases receptoras de lectina tipo L (LecRKs) na imunidade de plantas.
Consequentemente, a superexpressão de LecRK-I.9 aumentou a resistência a estes 3 patógenos. Este LecRK foi encontrado para associar com o receptor de flagelina FLS2, enfatizando sua especificidade na imunidade bacteriana [22]Estes dados indicam que os LecRKs têm um papel conservador na imunidade em todo o reino vegetal.
https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1006433
https://www.sciencedirect.com/science/article/pii/S0042682219300807
https://scholar.google.com.br/scholar?q=related:Nmc8nlHiifUJ:scholar.google.com/&scioq=&hl=pt-BR&as_sdt=2005&sciodt=0,5
O cabo-de-guerra entre plantas e vírus: grande progresso e muitas outras questões
X Wu, A Valli, JA Garcia, X Zhou, X Cheng - Vírus, 2019 - mdpi.com
As plantas são persistentemente desafiadas por vários fitopatógenos. Para se proteger, as plantas
evoluíram vigilância multicamadas contra todos os patógenos. Para parasitas intracelulares
vírus, as plantas desenvolveram imunidade inata, silenciamento de RNA, repressão da tradução ... https://www.mdpi.com/1999-4915/11/3/203
SGS3 Cooperates with RDR6 in Triggering Geminivirus-Induced Gene Silencing and in Suppressing Geminivirus Infection in Nicotiana Benthamiana J Mar Sci Eng, 2017 - https://www.mdpi.com/1999-4915/9/9/247?utm_source=TrendMD&utm_medium=cpc&utm_campaign=J_Mar_Sci_Eng_TrendMD_0
Which Plant Proteins Are Involved in Antiviral Defense? Review on In Vivo and In Vitro Activities of Selected Plant Proteins against Viruses Int J Mol Sci, 2017
A maioria dos vírus de plantas é vetorizada por insetos e as interações do vírus-planta-vetor têm importantes implicações ecológicas e evolutivas. Os vetores de insetos geralmente apresentam melhor desempenho em plantas infectadas por vírus. Esse mutualismo indireto entre vírus vegetais e insetos vetores promove a disseminação do vírus e tem efeitos agronômicos significativos. No entanto, poucos estudos investigaram como os vírus das plantas manipulam as defesas das plantas e promovem o desempenho dos vetores. Os begomovírus são um grupo proeminente de vírus de plantas em agroecossistemas tropicais e subtropicais e são transmitidos por moscas-brancas. Trabalhando com a mosca branca Bemisia tabaci, begomovírus e tabaco, nós revelamos que a proteína C2 de begomovírus sem satélites de DNA foi responsável pela supressão das defesas de plantas contra vetores de mosca branca. Descobrimos que a infecção de plantas pelo vírus enrolado em folhas amarelas de tomate (TYLCV), um dos begomovírus mais devastadores do mundo, promoveu a sobrevivência e reprodução de vetores de mosca branca. A proteína TYLCV C2 suprimiu as defesas das plantas ao interagir com a planta ubiquitina. Essa interação comprometeu a degradação da proteína JAZ1, inibindo a defesa do ácido jasmônico e a expressão dos genes da terpeno sintetase regulada por MYC2. Nós demonstramos ainda que a função da proteína C2 entre begomovírus não associados a satélites é bem conservada e a ubiquitinação é um alvo evolutivamente conservado de begomovírus para a supressão da resistência de plantas a vetores de mosca branca. Em conjunto, estes resultados demonstram que a inibição da ubiquitinação pela proteína C2 de begomovírus pode ser um mecanismo geral nas interações entre begomovírus, mosca-branca e planta. https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1007607&rev=1
C2 interage com o RPS27A in vitro e in vivo
Para elucidar como o TYLCV C2 manipula a defesa de plantas, nós selecionamos uma biblioteca de cDNA de tabaco por dois híbridos de levedura para identificar proteínas vegetais que interagem com C2. Nós projetamos um vetor de expressão de levedura contendo 1-78 aminoácido de C2 relatado como ausente do domínio de ativação transcricional [47]. Após o rastreio de dois híbridos de leveduras, foi descoberta uma proteína candidata N. tabacum RPS27A (NtRPS27A), que consiste num domínio de ubiquitina no terminal N e um domínio de proteína S27a ribossómica no terminal C, (Fig 3A). Com esta proteína candidata, primeiro confirmamos a interação entre C2 e NtRPS27A em dois híbridos de levedura (Fig. 3B). Em seguida, realizamos um ensaio de complementação de fluorescência bimolecular (BiFC) para examinar as interações in vivo de C2 e NtRPS27A. Fluorescência forte foi detectada em folhas nas quais NtRPS27A e C2 foram co-expressos (Fig. 3C). Em seguida, realizamos ensaios pull-down de GST e confirmamos a interação direta entre as duas proteínas de comprimento total (Fig 3D). O RPS27A contém dois domínios funcionais (Fig 3A). Para determinar qual domínio é responsável pela interação, realizamos ensaios BIFC e GST-pull down com RPS27A truncado. Nossos resultados mostraram que o domínio ubiquitina em vez do domínio RPS27a C-terminal (denominado RPS27Ac) poderia interagir com C2, indicando um papel da ubiquitina na interação (Fig 3C e 3D & S3 Fig). Como a proteína RPS27A triagem por dois híbridos de levedura é uma versão truncada contendo apenas 32-76 aminoácidos de NtRPS27A (ubiquitina32-76), especulamos que esse segmento possa determinar a interação entre C2 e NtRPS27A. Os ensaios de pull-down de BiFC e GST verificaram que a ubiquitina32-76 realmente interagia com C2 (Fig 3C e 3E). Além disso, detectamos a localização intracelular de C2 e diferentes segmentos de proteínas NtRPS27A usando a fusão GFP pelo sistema de expressão transiente. C2, NtRPS27A e ubiquitina estavam localizados no núcleo (Fig 3F), enquanto peptídeos contendo 32-76 aminoácidos da ubiquitina estavam localizados no núcleo e no citosol, semelhante à GFP livre (Fig. 3F). Considerados em conjunto, estes resultados confirmam que o C2 interage com o NtRPS27A, e os aminoácidos 32-76 do domínio da ubiquitina no NtRPS27A são importantes para a interação. Nós mostramos recentemente que a Rickettsia, um simbionte bacteriano facultativo secundário que infecta a mosca branca B. tabaci, está implicado na transmissão do vírus da folha amarela do tomate (TYLCV). A infecção com Rickettsia melhorou a aquisição e transmissão do vírus por adultos de B. tabaci. Aqui foi realizada uma análise transcriptômica com adultos de B. tabaci infectados e não infectados com Rickettsia, antes e após a aquisição do TYLCV. Os resultados mostram uma ativação dramática e específica do sistema imunológico na presença de Rickettsia antes da aquisição do TYLCV. No entanto, quando o TYLCV foi adquirido, ele induziu a ativação maciça da expressão gênica na população não infectada por Rickettsia, enquanto na população infectada por Rickettsia, o vírus induziu uma regulação negativa maciça da expressão gênica. Experimentos de condicionamento físico e escolha revelaram que, embora as moscas-brancas infectadas com Rickettsia sejam sempre mais atraídas por plantas infectadas com TYLCV, essa atração nem sempre é beneficiada por seus filhos. Esses estudos confirmam ainda mais o papel da Rickettsia em muitos aspectos das interações da B. tabaci com o TYLCV, e possivelmente servem como um fator importante na disseminação do vírus. https://europepmc.org/abstract/med/30762513
Os siRNAs intergênicos do vírus da onda da folha amarela do tomate têm como alvo um RNA não-codificante de longa permanência para modular os sintomas da doença
Y Yang, T Liu, D Shen, Wang J, X Ling, Z Hu… - PLoS…, 2019 - journals.plos.org
O vírus do enrolamento de folhas de tomate amarelo (TYLCV) e seus begomovírus relacionados causam disseminação rápida
doenças em tomate em todo o mundo. Como este vírus induz doenças ainda não está claro.
Aqui nós relatamos um modelo não codificado mediado por RNA para elucidar os mecanismos moleculares… https://journals.plos.org/plospathogens/article?id=10.1371/journal.ppat.1007534&rev=1